Le Heron et al. (2018) defines apathy as a marked reduction in goal-directed behavior. But in order to move, one must be motivated to do so. Therefore, a generalized form of impaired motivation also hallmarks apathy.
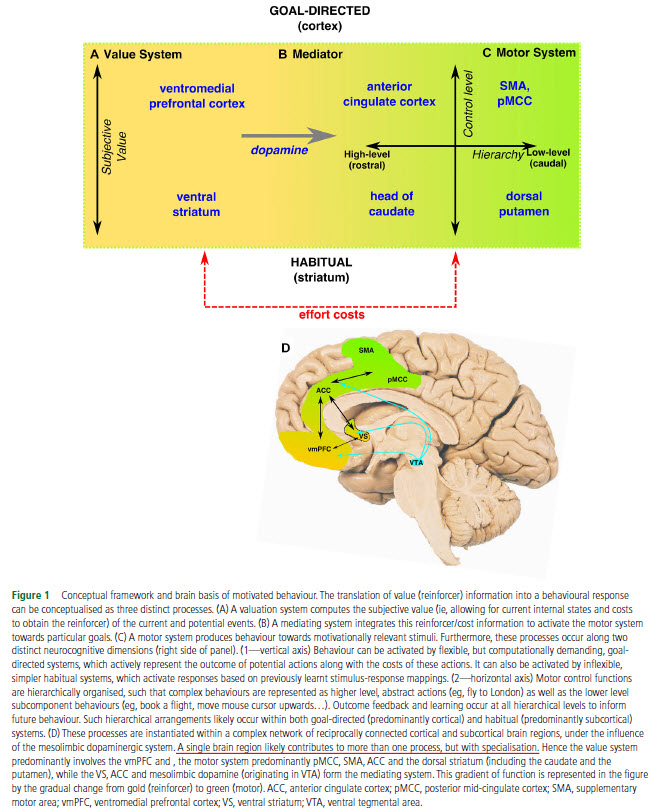
The authors compiled for us a nice mini-review combing through the literature of motivation in order to identify, if possible, the neurobiological mechanism(s) of apathy. First, they go very succinctly though the neuroscience of motivated behavior. Very succinctly, because there are literally hundreds of thousands of worthwhile pages out there on this subject. Although there are several other models proposed out-there, the authors’ new model on motivation includes the usual suspects (dopamine, striatum, prefrontal cortex, anterior cingulate cortex) and you can see it in the Fig. 1.
After this intro, the authors go on to showcasing findings from the effort-based decision-making field, which suggest that the dopamine-producing neurons from ventral tegmental area (VTA) are fundamental in choosing an action that requires high-effort for high-reward versus a low-effort for low-reward. Contrary to what Wikipedia tells you, a reduction, not an increase, in mesolimbic dopamine is associated with apathy, i.e. preferring a low-effort for low-reward activity.
Next, the authors focus on why are the apathetic… apathetic? Basically, they asked the question: “For the apathetic, is the reward too little or is the effort too high?” By looking at some cleverly designed experiments destined to parse out sensitivity to reward versus sensitivity to effort costs, the authors conclude that the apathetics are indeed sensitive to the reward, meaning they don’t find the rewards good enough for them to move. Therefore, the answer is the reward is too little.
In a nutshell, apathetic people think “It’s not worth it, so I’m not willing to put in the effort to get it”. But if somehow they are made to judge the reward as good enough, to think “it’s worth it”, they are willing to work their darndest to get it, like everybody else.
The application of this is that in order to get people off the couch and do stuff you have to present them a reward that they consider worth moving for, in other words to motivate them. To which any practicing psychologist or counselor would say: “Duh! We’ve been saying that for ages. Glad that neuroscience finally caught up”. Because it’s easy to say people need to get motivated, but much much harder to figure out how.
This was a difficult write for me and even I recognize the quality of this blogpost as crappy. That’s because, more or less, this paper is within my narrow specialization field. There are points where I disagree with the authors (some definitions of terms), there are points where things are way more nuanced than presented (dopamine findings in reward), and finally there are personal preferences (the interpretation of data from Parkinson’s disease studies). Plus, Salamone (the second-to-last author) is a big name in dopamine research, meaning I’m familiar with his past 20 years or so worth of publications, so I can infer certain salient implications (one dopamine hypothesis is about saliency, get it?).
It’s an interesting paper, but it’s definitely written for the specialist. Hurray (or boo, whatever would be your preference) for another model of dopamine function(s).
Although a complete musical anti-talent myself, that doesn’t prohibit me from fully enjoying the works of the masters in the art. When my family is out of earshot, I even bellow – because it cannot be called music – from the top of my lungs alongside the most famous tenors ever recorded. A couple of days ago I loaded one of my most eclectic playlists. While remembering my younger days as an Iron Maiden concert goer (I never said I listen only to classical music :D) and screaming the “Fear of the Dark” chorus, I wondered what’s new on the front of music processing in the brain.
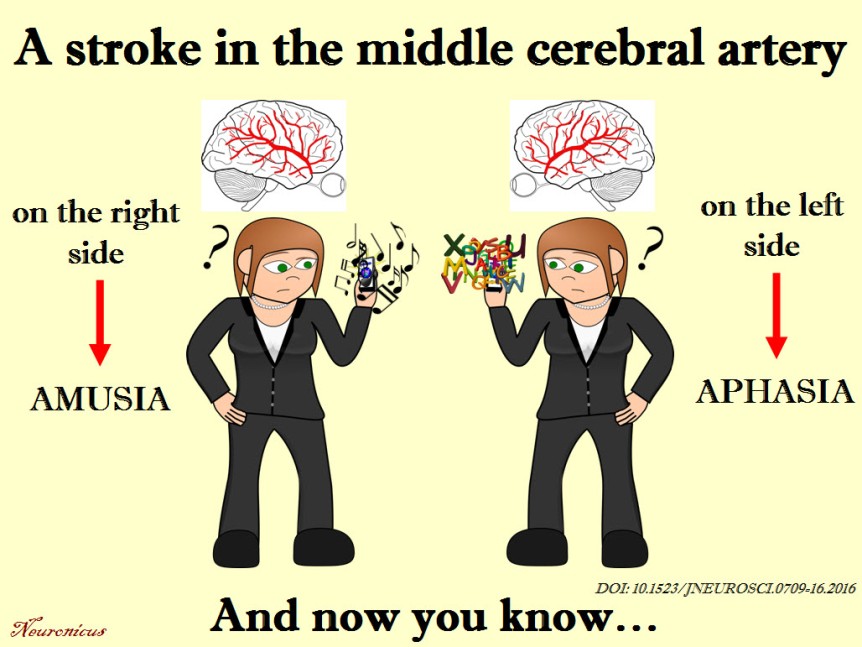
And I found an interesting recent paper about amusia. Amusia is, as those of you with ancient Greek proclivities might have surmised, a deficit in the perception of music, mainly the pitch but sometimes rhythm and other aspects of music. A small percentage of the population is born with it, but a whooping 35 to 69% of stroke survivors exhibit the disorder.
So Sihvonen et al. (2016) decided to take a closer look at this phenomenon with the help of 77 stroke patients. These patients had an MRI scan within the first 3 weeks following stroke and another one 6 months poststroke. They also completed a behavioral test for amusia within the first 3 weeks following stroke and again 3 months later. For reasons undisclosed, and thus raising my eyebrows, the behavioral assessment was not performed at 6 months poststroke, nor an MRI at the 3 months follow-up. It would be nice to have had behavioral assessment with brain images at the same time because a lot can happen in weeks, let alone months after a stroke.
Nevertheless, the authors used a novel way to look at the brain pictures, called voxel-based lesion-symptom mapping (VLSM). Well, is not really novel, it’s been around for 15 years or so. Basically, to ascertain the function of a brain region, researchers either get people with a specific brain lesion and then look for a behavioral deficit or get a symptom and then they look for a brain lesion. Both approaches have distinct advantages but also disadvantages (see Bates et al., 2003). To overcome the disadvantages of these methods, enter the scene VLSM, which is a mathematical/statistical gimmick that allows you to explore the relationship between brain and function without forming preconceived ideas, i.e. without forcing dichotomous categories. They also looked at voxel-based morphometry (VBM), which a fancy way of saying they looked to see if the grey and white matter differ over time in the brains of their subjects.
After much analyses, Sihvonen et al. (2016) conclude that the damage to the right hemisphere is more likely conducive to amusia, as opposed to aphasia which is due mainly to damage to the left hemisphere. More specifically,
“damage to the right temporal areas, insula, and putamen forms the crucial neural substrate for acquired amusia after stroke. Persistent amusia is associated with further [grey matter] atrophy in the right superior temporal gyrus (STG) and middle temporal gyrus (MTG), locating more anteriorly for rhythm amusia and more posteriorly for pitch amusia.”
The more we know, the better chances we have to improve treatments for people.
unless you’re left-handed, then things are reversed.
Among the many humorous sayings, puns, and jokes that one inevitably encounters on any social medium account, one that was popular this year was about the similarity between putting a 2 year old to bed and putting your drunk friend to bed, which went like this: they both sing to themselves, request water, mumble and blabber incoherently, do some weird yoga posses, cry, hiccup, and then they pass out. The joke manages to steal a smile only if someone has been through both situations, otherwise it looses its appeal.
Being exposed to both situations, I thought that while the water request from the drunk friend is a response to the dehydrating effects of alcohol, the water request from the toddler is probably nothing more than a delaying tactic to postpone bedtime. Whether there may or may not be some truth to my assumption in the case of the toddler, here is a paper to show that there is definitely more to the water request than meets the eye.
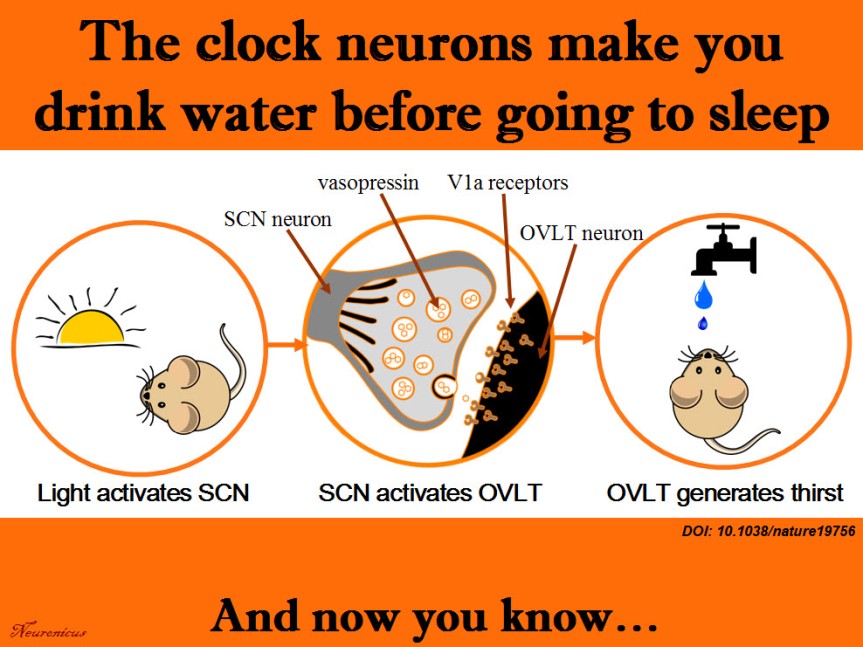
Generally, thirst is generated by the hypothalamus when its neurons and neurons from organum vasculosum lamina terminalis (OVLT) in the brainstem sense that the blood is either too viscous (hypovolaemia) or too salty (hyperosmolality), both phenomena indicating a need for water. Ingesting water would bring these indices to homeostatic values.
More than a decade ago, researchers observed that rodents get a good gulp of water just before going to sleep. This surge was not motivated by thirst because the mice were not feverish, were not hungry and they did not have a too viscous or a too salty blood. So why do it then? If the rodents are restricted from drinking the water they get dehydrated, so obviously the behavior has function. But is not motivated by thirst, at least not the way we know it. Huh… The authors call this “anticipatory thirst”, because it keeps the animal from becoming dehydrated later on.
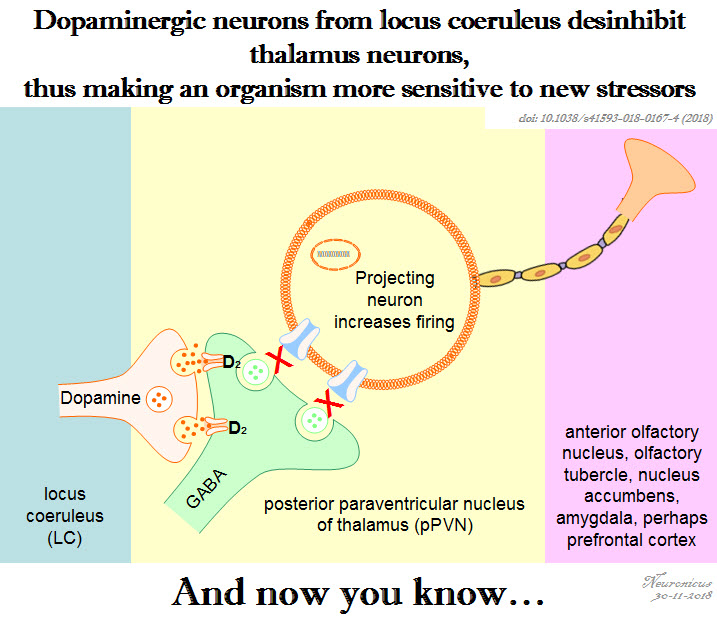
Since the behavior occurs with regularity, maybe the neurons that control circadian rhythms have something to do with it. So Gizowski et al. (2016) took a closer look at the activity of clock neurons from the suprachiasmatic nucleus (SCN), a well known hypothalamic nucleus heavily involved in circadian rhythms. The authors did a lot of work on SCN and OVLT neurons: fluorescent labeling, c-fos expression, anatomical tracing, optogenetics, genetic knockouts, pharmacological manipulations, electrophysiological recordings, and behavioral experiments. All these to come to this conclusion:
SCN neurons release vasopressin and that excites the OVLT neurons via V1a receptors. This is necessary and sufficient to make the animal drink the water, even if it’s not thirsty.
That’s a lot of techniques used in a lot of experiments for only three authors. Ten years ago, you needed only one, maybe two techniques to prove the same point. Either there have been a lot of students and technicians who did not get credit (there isn’t even an Acknowledgements section. EDIT: yes, there is, see the comments below or, if they’re missing, the P.S.) or these three authors are experts in all these techniques. In this day and age, I wouldn’t be surprised by either option. No wonder small universities have difficulty publishing in Big Name journals; they don’t have the resources to compete. And without publishing, no tenure… And without tenure, less research… And thus shall the gap widen.
Musings about workload aside, this is a great paper, shedding light on yet another mysterious behavior and elucidating the mechanism behind it. There’s still work to be done though, like answering how accurate is the SCN in predicting bedtime to activate the drinking behavior. Does it take its cues from light only? Does ambient temperature play a role and so on. This line of work can help people that work in shifts to prevent certain health problems. Their SCN is out of rhythm and that can influence deleteriously the activity of a whole slew of organs.
Summary of the doi: 10.1038/nature19756 findings. 1) The light is a cue for suprachiasmatic nulceus (SCN) that bedtime is near. 2) The SCN vasopressin neurons that project to organum vasculosum lamina terminalis (OVLT) are activated. 3) The OVLT generates the anticipatory thirst. 4) The animal drinks fluids.
EDIT (12 Oct 2016): P.S. The blog comments are automatically deleted after a period of time. In case of this post that would be a pity because I have been fortunate to receive comments from at least one of the authors of the paper, the PI, Dr. Charles Bourque and, presumably under pseudonym, but I don’t know that for sure, also the first author, Claire Gizowski. So I will include here, in a post scriptum, the main idea of their comments. Here is an excerpt from Dr. Bourque’s comment:
“Let me state for the record that Claire accomplished pretty much ALL of the work in this paper (there is a description of who did what at the end of the paper). More importantly, there were no “unthanked” undergraduates, volunteers or other parties that contributed to this work.”
My hat, Ms. Gizowski. It is tipped. To you. Congratulations! With such an impressive work I am sure I will hear about you again and that pretty soon I will blog about Dr. Gizowski.
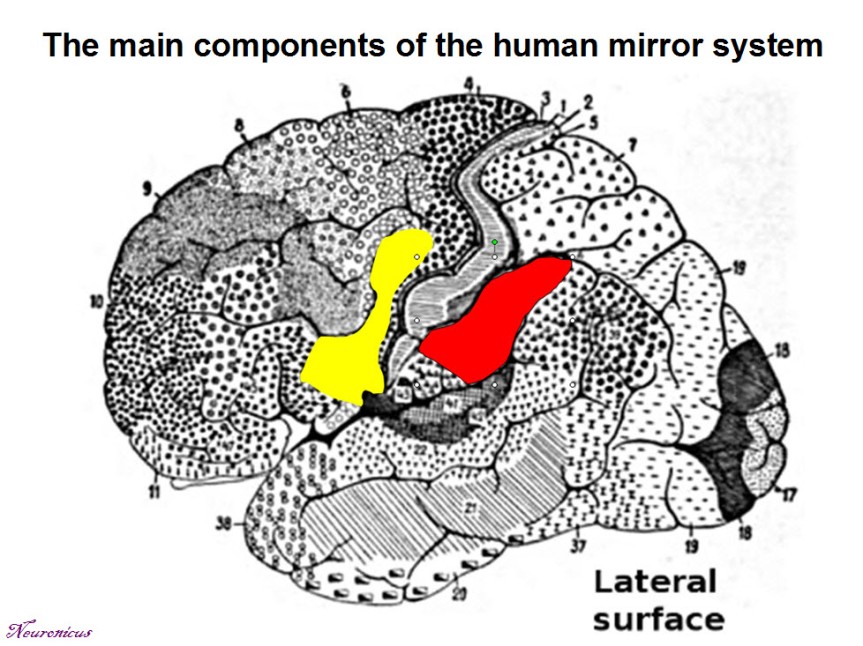
There are some neurons in the human brain that fire both when the person is doing some behavior and when watching that behavior performed by someone else. These cells are called mirror neurons and were first discovered in 1988 (see NOTE) by a group of researchers form the University of Parma, Italy, led by Giacomo Rizzolatti.
The discovery was done by accident. The researchers were investigating the activity of neurons in the rostral part of the inferior premotor cortex (riPM) of macaque monkeys with electrophysiological recordings. They placed a box in front of the monkey which had various objects in it. When the monkey pressed a switch, the content of the box was illuminated, then a door would open and the monkey reached for an object. Under each object was hidden a small piece of food. Several neurons were discharging when the animal was grasping the object. But the researchers noticed that some of these neurons ALSO fired when the monkey was motionless and watching the researcher grasping the objects!
The authors then did more motions to see when exactly the two neurons were firing, whether it’s related to the food or threatening gestures and so on. And then they recorded from some 182 more neurons while the monkey or the experimenter were performing hand actions with different objects. Importantly, they also did an electromyogram (EMG) and saw that when the neurons that were firing when the monkey was observing actions, the muscles did not move at all.
They found that some neurons responded to both when doing and seeing the actions, whereas some other neurons responded only when doing or only when seeing the actions. The neurons that are active when observing are called mirror neurons now. In 1996 they were identified also in humans with the help of positron emission tomography (PET).
It is tragicomical that the authors first submitted their findings to the most prestigious scientific journal, Nature, believing that their discovery is worth it, and rightfully so. But, Nature rejected their paper because of its “lack of general interest” (Rizzolatti & Fabbri-Destro, 2010)! Luckily for us, the editor of Experimental Brain Research, Otto Creutzfeld, did not share Nature‘s opinion.
Thousands of experiments followed the tremendous discovery of mirror neurons, even trying to manipulate their activity. Many researchers believe that the activity of the mirror neurons is fundamental for understanding the intentions of others, the development of theory of mind, empathy, the process of socialization, language development and even human self-awareness.
NOTE: Whenever possible, I try to report both the date of the discovery and the date of publication. Sometimes, the two dates can differ quite a bit. In this case, the discovery was done in 1988 and the publishing in 1992.
References:
di Pellegrino G, Fadiga L, Fogassi L, Gallese V, & Rizzolatti G (October 1992). Understanding motor events: a neurophysiological study. Experimental Brain Research, 91(1):176-180. DOI: 10.1007/BF00230027. ARTICLE | Research Gate FULLTEXT PDF
Rizzolatti G & Fabbri-Destro M (Epub 18 Sept 2009; January 2010). Mirror neurons: From discovery to autism. Experimental Brain Research, 200(3): 223-237. DOI: 10.1007/s00221-009-2002-3.ARTICLE | Research Gate FULLTEXT PDF
A few decades ago, Italian researchers from the University of Parma discovered some neurons in monkey which were active not only when the monkey is performing an action, but also when watching the same action performed by someone else. This kind of neuron, or rather this particular neuronal behavior, had been subsequently identified in humans scattered mainly within the frontal and parietal cortices (front and top of your head) and called the mirror neuron system (MNS). Its role is to understand the intentions of others and thus facilitate learning. Mind you, there are, as it should be in any healthy vigorous scientific endeavor, those who challenge this role and even the existence of MNS.
Hobson & Bishop (2016) do not question the existence of the mirror neurons or their roles, but something else. You see, proper understanding of intentions, actions and emotions of others is severely impaired in autism or some schizophrenias. Correspondingly, there have been reports saying that the MNS function is abnormal in these disorders. So if we can manipulate the neurons that help us understanding others, then we may be able to study the neurons better, and – who knows? – maybe even ‘switch them on’ and ‘off’ when needed (Ha! That’s a scary thought!).
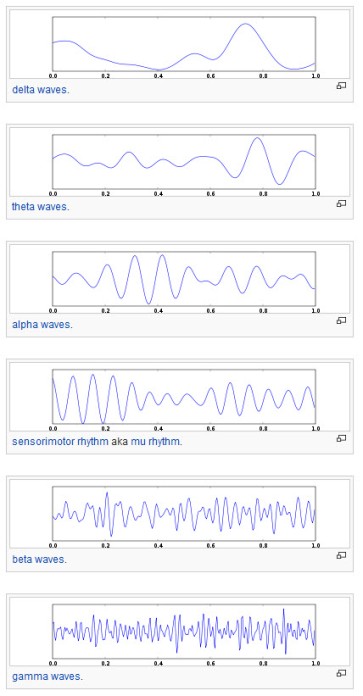
Human EEG waves (from Wikipedia, under CC BY-SA 3.0 license)
Anyway, previous work said that recording a weak Mu frequency in the brain regions with mirror neurons show that these neurons are active. This frequency (between 8-13 Hz) is recorded through electroencephalography (EEG). The assumption is as follows: when resting, neurons fire synchronously; when busy, they fire each to its own, so they desynchronize, which leads to a reduction in the Mu intensity.
All well and good, but there is a problem. There is another frequency that overlaps with the Mu frequency and that is the Alpha band. Alpha activity is highest when a person is awake with eyes closed, but diminishes when the person is drowsy or, importantly, when making a mental effort, like paying great attention to something. So, if I see a weak Mu/Alpha frequency when the subject is watching someone grabbing a pencil, is that because the mirror neurons are active or because he’s sleepy? There are a few gimmicks to disentangle between the two, from the setup of the experiment in such a way that it requires same attention demand over tasks to the careful localization of the origin of the two waves (Mu is said to arise from sensoriomotor regions, whereas Alpha comes from more posterior regions).
But Hobson & Bishop (2016) argue that this disentangling is more difficult than previously thought by carrying out a series of experiments where they varied the baseline, in such a way that some were more attentionally demanding than others. After carefully analyzing various EEG waves and electrodes positions in these conditions, they conclude that “mu suppression can be used to index the human MNS, but the effect is weak and unreliable and easily confounded with alpha suppression“.
What makes this paper interesting to me, besides its empirical findings, is the way the experiment was conducted and published. This is a true hypothesis driven study, following the scientific method step by step, a credit to us all scientists. In other words, a rare gem. A lot of other papers are trying to make a pretty story from crappy data or weave some story about the results as if that’s what they went for all along when in fact they did a bunch of stuff and chose what looked good on paper.
Let me explain. As a consequence of the incredible pressure put on researchers to publish or perish (which, believe me, is more than just a metaphor, your livelihood and career depend on it), there is an alarming increase in bad papers, which means
papers with inappropriate statistical analyses (p threshold curse, lack of multiple comparisons corrections, like the one brilliantly exposed here),
papers with huge databases in which some correlations are bound to appear by chance alone and are presented as meaningful (p-hacking or data fishing),
papers without enough data to make a meaningful conclusion (lack of statistical power),
papers that report only good-looking results (only positive results required by journals),
papers that seek only to provide data to reinforce previously held beliefs (confirmation bias)
and so on.
For these reasons (and more), there is a high rate of rejection of papers submitted to journals (about 90%), which means more than just a lack of publication in a good journal; it means wasted time, money and resources, shattered career prospects for the grad students who did the experiments and threatened job security for everybody involved, not to mention a promotion of distrust of science and a disservice to the scientific endeavor in general. So some journals, like Cortex, are moving toward a system called Registered Report, which asks for the rationale and the plan of the experiment before this is conducted, which should protect against many of the above-mentioned plagues. If the plan is approved, the chances to get the results published in that journal are 90%.
This is one of those Registered Report papers. Good for you, Hobson & Bishop!
REFERENCE: Hobson HM & Bishop DVM (Epub April 2016). Mu suppression – A good measure of the human mirror neuron system?. Cortex, doi: 10.1016/j.cortex.2016.03.019 ARTICLE | FREE FULLTEXT PDF | RAW DATA
Nothing short of an autism cure is promised by this hot new research paper.
Among many thousands of proteins that a neuron needs to make in order to function properly there is one called SHANK3 made from the gene shank3. (Note the customary writing: by consensus, a gene’s name is written using small caps and italicized, whereas the protein’s name that results from that gene expression is written with caps).
This protein is important for the correct assembly of synapses and previous work has shown that if you delete its gene in mice they show autistic-like behavior. Similarly, some people with autism, but by far not all, have a deletion on Chromosome 22, where the protein’s gene is located.
The straightforward approach would be to restore the protein production into the adult autistic mouse and see what happens. Well, one problem with that is keeping the concentration of the protein at the optimum level, because if the mouse makes too much of it, then the mouse develops ADHD and bipolar.
So the researchers developed a really neat genetic model in which they managed to turn on and off the shank3 gene at will by giving the mouse a drug called tamoxifen (don’t take this drug for autism! Beside the fact that is not going to work because you’re not a genetically engineered mouse with a Cre-dependent genetic switch on your shank3, it is also very toxic and used only in some form of cancers when is believed that the benefits outweigh the horrible side effects).
In young adult mice, the turning on of the gene resulted in normalization of synapses in the striatum, a brain region heavily involved in autistic behaviors. The synapses were comparable to normal synapses in some aspects (from the looks, i.e. postsynaptic density scaffolding, to the works, i.e. electrophysiological properties) and even more so in others (more dendritic spines than normal, meaning more synapses, presumably). This molecular repair has been mirrored by some behavioral rescue: although these mice still had more anxiety and more coordination problems than the control mice, their social aversion and repetitive behaviors disappeared. And the really really cool part of all this is that this reversal of autistic behaviors was done in ADULT mice.
Now, when the researchers turned the gene on in 20 days old mice (which is, roughly, the equivalent of the entering the toddling stage in humans), all four behaviors were rescued: social aversion, repetitive, coordination, and anxiety. Which tells us two things: first, the younger you intervene, the more improvements you get and, second and equally important, in adult, while some circuits seem to be irreversibly developed in a certain way, some other neural pathways are still plastic enough as to be amenable to change.
Awesome, awesome, awesome. Even if only a very small portion of people with autism have this genetic problem (about 1%), even if autism spectrum disorders encompass such a variety of behavioral abnormalities, this research may spark hope for a whole range of targeted gene therapies.
Reference: Mei Y, Monteiro P, Zhou Y, Kim JA, Gao X, Fu Z, Feng G. (Epub 17 Feb 2016). Adult restoration of Shank3 expression rescues selective autistic-like phenotypes. Nature. doi: 10.1038/nature16971. Article | MIT press release
Most of the time the brain receives information from different senses about the same object or event. For example, to localize an object that makes noise we use both visual and auditory information – if these are available to us -, process called multisensory information integration (MSII).
It is generally believed that the way this integration happens in a physical sense is by getting all these data in a special brain region dedicated to integrate information from multiple senses. And that there are several regions like that in the brain; for example superior colliculi integrate the visual and auditory information from the example above. This belief is not without empirical support. Indeed, many experiments both in vivo (i.e. in the awake behaving animal) and in silico (i.e. simulated on a computer program by building neural networks) have strengthened this idea.
But Zhang et al. (2016) claim there is some data that doesn’t fit the model. So let’s build a different model. Which is exactly what they have done using continuous attractor neural networks (CANN) as the building blocks for a neural network that seems to be biologically realistic. The output of their experiments shows that instead of having a central processing area in the brain that integrates data from multiple senses, optimal processing can be achieved by a decentralized network with neurons that are reciprocally connected.
Today, uncharacteristically, I am covering a paper that I have not read in its entirety. That is because, frankly, they lost me after the first Methods paragraph where they describe the way they built the neural network. In my defense, I don’t think there are many people in the world outside the neural networks field that can follow the mathematical formulae (see the Excerpt pic).
Excerpt from the Methods section of Zhang et al. (2016, doi: 10.1523/JNEUROSCI.0578-15.2016.)
Anyway, my gut instinct is that both hypotheses have merit in that the brain uses both specialized multisensory areas, like superior colliculi and decentralized, distributed reciprocal connections, like the model proposed by Zhang et al. (2016).
Reference: Zhang WH, Chen A, Rasch MJ, & Wu S (13 Jan 2016). Decentralized Multisensory Information Integration in Neural Systems. Journal of Neuroscience, 36(2):532-47. doi: 10.1523/JNEUROSCI.0578-15.2016.Article | FREE Fulltext PDF
When you’re sick you also feel awful: no appetite, weak, sleepy, feverish, achy, and so on. This is called, appropriately so, the sickness syndrome.
Saper, Romanovsky & Scammell (2012) wrote a beautiful review of the neural circuits underlying this collection of symptoms. In a nutshell, the immune system releases cytokines to fight the inflammation, which in turn stimulate the release of prostaglandins. Prostaglandins bind to various areas in the brain to produce the sickness syndrome symptoms. Below are outlined four simplified brain circuits which the non-specialists can skip entirely.
Prostaglandins in the median preoptic nucleus lead to a cascade involving dorsomedial hypothalamus, rostral medullary raphe and finally the spinal cord to produce fever by activating the brown adipose tissue.
Prostaglandins in the preoptic area lead to the inhibition of the brain’s analgesic system involving the descending projections of the periaqueductal grey to spinal cord, thus promoting achiness.
Prostaglandins in the meninges result in adenosine release in nucleus accumbens and ventrolateral preoptic nucleus which, downstream, end in inhibiting the arousal system to produce sleepiness.
Prostaglandins in the arcuate nucleus lead to inhibition of several hypothalamic nuclei involved in promoting feeding, thereby producing anorexia.
The sickness syndrome and the role prostaglandins play in it has tremendous adaptive role, as it promotes rest and recuperation. So don’t blame them too much. And if you’re really done feeling sick, take some non-steroid anti-inflammatory drugs, like aspirin, which inhibit the prostaglandins’ synthesis very effectively. That’s how and why NSAIDs work.
Reference: Saper CB, Romanovsky AA & Scammell TE (26 Jul 2012). Neural Circuitry Engaged by Prostaglandins during the Sickness Syndrome.Nature Neuroscience, 15(8):1088-95. doi: 10.1038/nn.3159. Article | FREE Fulltext PDF
Vagus nerve stimulation by a wireless device implanted subcutaneously. License: PD. Credit: NIH/NIMH
A stroke is a serious medical condition that is characterized by the death of brain cells following bleeding in the brain or lack of blood supply to those cells. Three quarters of the survivors have weakness in the arm which can be permanent. Physical therapy helps, but not much.
Dawson et al. (2015) report a novel way to increase arm mobility after stroke. Previous findings in animal studies showed promising results by stimulating the vagus nerve (VNS). This nerve is the tenth out of the twelve cranial nerves and has many functions, primarily heart and digestive control. The authors implanted a small wireless device in the neck of nine stroke survivors and delivered very short (half a second) mild (0.8 mA) electrical pulses during rehabilitation therapy. Ten matched controls received rehab therapy only.
The authors measured motor recovery by several tests, one of which is Fugl–Meyer assessment-UE. At this test, the rehab only group improved by 3 points and the VNS + rehab group by about 9 points and this difference was statistically significant (I believe this scale’s upper limit is 66, but I’m not 100% sure).
Although the authors offer a possible mechanism through which VNS might produce cortical plasticity (through release of acetylcholine and norepinephrine driven by activation of nucleus tractus solitarius) the truth is that we don’t really know how it works. Nevertheless, it seems that VNS paired with rehab is better than rehab alone, and that in itself certainly warrants further studies, perhaps the next step being a fully double-blind experiment.
Reference: Dawson J, Pierce D, Dixit A, Kimberley TJ, Robertson M, Tarver B, Hilmi O, McLean J, Forbes K, Kilgard MP, Rennaker RL, Cramer SC, Walters M, & Engineer N. (Epub 8 Dec 2015). Safety, Feasibility, and Efficacy of Vagus Nerve Stimulation Paired With Upper-Limb Rehabilitation After Ischemic Stroke. Stroke. 2016; 47:00-00. DOI: 10.1161/STROKEAHA.115.010477. Article| FREE FULLTEXT PDF
A woman named BT suffered an accident when she was 20 years old and she became blind. Thirteen year later she was referred to Bruno Waldvogel (one of the two authors of the paper) for psychotherapy by a psychiatry clinic who diagnosed her with dissociative identity disorder, formerly known as multiple personality disorder.
The cortical blindness diagnosis has been established after extensive ophtalmologic tests in which she appeared blind but not because of damage to the eyes. So, by inference, it had to be damage to the brain. Remarkably (we shall see later why), she had no oculomotor reflexes in response to glare. Moreover, visual evoked potentials (VEP is an EEG in the occipital region) showed no activity in the primary visual area of the brain (V1).
During the four years of psychotherapy, BT showed more than 10 distinct personalities. One of them, a teenage male, started to see words on a magazine and pretty soon could see everything. With the help of hypnotherapeutic techniques, more and more personalities started to see.
“Sighted and blind states could alternate within seconds” (Strasburger & Waldvogel, 2015).
The VEP showed no or very little activity when the blind personality was “on” and showed normal activity when the sighted personality was “on”. Which is extremely curious, because similar studies in people with psychogenic blindness or anesthetized showed intact VEPs.
There are a couple of conclusions from this: 1) BT was misdiagnosed, as is unlikely to be any brain damage because some personalities could see, and 2) Multiple personalities – or dissociate identities, as they are now called – are real in the sense that they can be separated at the biological level.
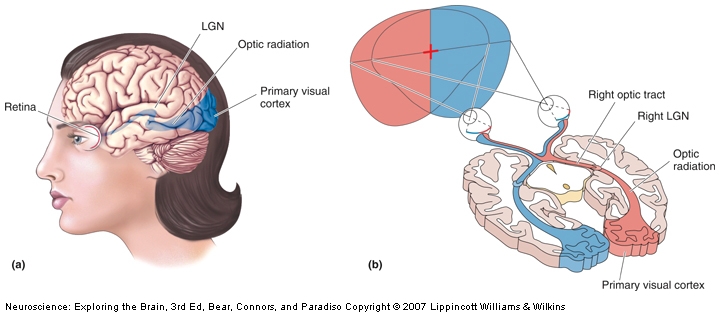
The visual pathway that mediates conscious visual perception. a) A side view of the human brain with the retinogeniculocortical pathway shown inside (blue). b) A horizontal section through the brain exposing the same pathway.
Fascinating! The next question is, obviously, what’s the mechanism behind this? The authors say that it’s very likely the LGN (the lateral geniculate nucleus of the thalamus) which is the only relay between retina and V1 (see pic). It can be. Surely is possible. Unfortunately, so are other putative mechanisms, as 10% of the neurons in the retina also go to the superior colliculus, and some others go directly to the hypothalamus, completely bypassing the thalamus. Also, because it is impossible to have a precise timing on the switching between personalities, even if you MRI the woman it would be difficult to establish if the switching to blindness mode is the result of a bottom-up or a top-down modulation (i.e. the visual information never reaches V1, it reaches V1 and is suppressed there, or some signal form other brain areas inhibits V1 completely, so is unresponsive when the visual information arrives).
Despite the limitations, I would certainly try to get the woman into an fMRI. C’mon, people, this is an extraordinary subject and if she gave permission for the case study report, surely she would not object to the scanning.
Reference: Strasburger H & Waldvogel B (Epub 15 Oct 2015). Sight and blindness in the same person: Gating in the visual system. PsyCh Journal. doi: 10.1002/pchj.109.Article | FULLTEXT PDF|Washington Post cover
The runner’s high is most likely due to release of the endorphins binding to the opioid receptors according to Boecker et al. (2008, doi: 10.1093/cercor/bhn013). Image courtesy of Pixabay.
We all know that exercise is good for you: it keeps you fit, it reduces stress and improves your mood. And also, sometimes, particularly after endurance running, it gets you high. The mechanism of euphoria reported by some runners after resistance training is unknown. Here is a nice paper trying to figure it out.
Boecker et al. (2008) scanned 10 trained male athletes at rest and after 2 hour worth of endurance running. By “trained athletes” they mean people that ran for 4-10 hours weekly for the past 2 years. The scanning was done using a positron emission tomograph (PET). The PET looks for a particular chemical that has been injected into the bloodstream of the subjects, in this case non-selective opioidergic ligand (it binds to all opioid receptors in the brain; morphine, for example, binds only to a subclass of the opioid receptors).
The rationale is as follows: if we see an increase in ligand binding, then the receptors were free, unoccupied, showing a reduction in the endogenous neurotransmitter, that is the substance that the brain produces for those receptors; if we see a decrease in the ligand binding it was because the receptors were occupied, meaning that there was an increase in the production of the endogenous neurotransmitter. The endogenous neurotransmitters for the opioid receptors are the endorphins (don’t confuse them with epinephrine a.k.a. adrenaline; different systems entirely).
After running, the subjects reported that they are euphoric and happy, but no change in other feelings (confusion, anger, sadness, fear etc.; there was a reduction in fear, but it was not significant). The scanning showed that it was less binding of the opioidergic ligand in many places in the brain (for the specialist, here you go: prefrontal/orbitofrontal cortices, dorsolateral prefrontal cortex, anterior and posterior cingulate cortex, insula and parahippocampal gyrus, sensorimotor/parietal regions, cerebellum and basal ganglia).
Regression analysis showed that there was a link between the euphoria feeling and the receptor occupancy: the more euphoric the people said they were, the more endorphines (i.e. endogenous opioids) they had bound in the brain. This study is the first to show this kind of link.
Reference: Boecker H, Sprenger T, Spilker ME, Henriksen G, Koppenhoefer M, Wagner KJ, Valet M, Berthele A, & Tolle TR. (Nov 2008, Epub 21 Feb 2008). The Runner’s High: Opioidergic Mechanisms in the Human Brain. Cerebral Cortex, 18:2523–2531. doi:10.1093/cercor/bhn013. Article | FREE FULLTEXT PDF
The pons varolii, as described in plate 677 of Henry Gray’s Anatomy of the Human Body (1918). License: PD.
The name of the pons, that part of the brainstem that is so important for survival functions (like breathing) and holds the nuclei of several cranial nerves, is actually pons varolii. I was wondering why is that? When I learned neuroanatomy I was extremely lucky, because my knowledge of Latin, such as it is, contributed immensely to the memorization of brain structures; so the name of pons means “bridge” in Latin, which makes sense because it looks like one (see picture). But I was at a loss with varolii. Was it some sort of a joke that I missed? Was it the “rude bridge” or, more colloquially, the “a**hole bridge”?! Varo (or the closest thing) in Latin means rude or uncivilized.
Title page of Varolio’s published letter in 1573.
Well, turns out that the guy who described the pons for the first time is Costanzo Varolio (1543–1575) and the structure is named after him. Duh! As if it’s uncommon to name things after their discoverer… Anyway, I didn’t read the original account, which is free in its digitized-by-Google form of dubious quality (you can see the actual thumb of the dude who scanned it on the last page and many pages are illegible due to poor scanning technique). I got the information about the pons from the Pioneers in Neurology section in the Journal of Neurology. Varolio wrote a huge letter (seventy-some pages worth!) on 1 April 1572 to another physician describing the optical nerves and the pons. The letter has been published a year later in Padua, Italy. The pons may have been described and/or named earlier, but, alas, the works were not published or published much later. Goes to show that publication is more important that discovery…
Original reference: Varolio, C. (1573). De Nervis Opticis nonnullisque aliis praeter communem opinionem in Humano capite observatis (On the optic nerves observed in the human brain and a few other particulars adverse to the common opinion). Padua. Google ebook
Reference: Zago S & Meraviglia MV (July 2009, Epub 6 June 2009). Costanzo Varolio (1543–1575). Journal of Neurology, 256(7):1195-6. doi: 10.1007/s00415-009-5192-5. Article | FREE FULLTEXT PDF
Contrary to popular belief, rats and mice are very fastidious animals; they keep themselves scrupulously clean by engaging in a very meticulous routine of self-grooming. The routine is so rigorous that allows the researchers to divide the grooming sequence into four different phases, starting with the nose and whiskers and ending with the genitalia and tail. It is also a symmetrical behavior (no whisker left ungroomed, no paw unlicked).
Grooming is sensitive to dopaminergic manipulations, so Pelosi, Girault, & Hervé (2015) sought to see what happens if they destroy the dopamine fibers in the mouse brain. So they lesioned the medial forebrain bundle, which is a bunch axon fibers that contains over 80% of the midbrain dopaminergic axons. But they were tricky, they lesioned only one side.
And the results were that the lesioned mice not only exhibited less self-grooming on the opposite side to the lesion, but the behavior was rescued by L-DOPA, which is medication for Parkinson’s. That is, they gave the mice some L-DOPA and they began to merrily self-groom again on both sides of the body. The authors discuss in depths other findings, like the changes (or absence thereof) in grooming bouts, grooming time, grooming bouts, completeness of grooming etc.
The findings have significance in the Parkinson’s research, where the mild to moderate phases of the disease often present with asymmetrical motor behavior.
Reference: Pelosi A, Girault J-A, & Hervé D (23 Sept 2015). Unilateral Lesion of Dopamine Neurons Induces Grooming Asymmetry in the Mouse. PLoS One. 2015; 10(9): e0137185. doi: 10.1371/journal.pone.0137185. PMCID: PMC4580614. Article | FREE FULLTEXT PDF
The nucleus accumbens, a key region involved in reward processing and addiction. Credit: Zou et al. (2015)
Consumption of addictive drugs changes your brain and these changes underlie the consequent dependence. It is very difficult to quit, and certainly it is not a matter of lack of will power that the majority of drug users have such a great difficulty in trying to quit. But what happens to the brains of the lucky few who managed to kick the addiction out the window? Are their brains reverting to their pre-addiction states? As the paper below shows, sadly, no. But there is hope.
Zhou et al. ( 2015) used resting state fMRI (which means scanning your brain without requiring you to perform any task) to investigate the brains of 30 healthy controls and 30 heroin-addicts who were abstinent from the drug for more than 3 years. Specifically, they looked to see if the connectivity of certain brain regions involved in addiction is different after such a long abstinence time.
The bad news is that the abstinents still had some abnormal connectivity (for specialists: “stronger functional connectivity between the nucleus accumbens and the right ventromedial prefrontal cortex and relatively weaker connectivity between the nucleus accumbens and the left putamen, left precuneus, and supplementary motor area” p. 1697).
The good news: the longer the abstinence time, the greater the strength of the connection between nucleus accumbens and putamen (first structure involved in reward processing, the second in habit-learning), suggesting a partial neural recovery.
The study has some limitations: technical (did not control for heartbeat and respiration), methodological (most of the heroin abstinents were smokers, another addiction) and theoretical (one brain area does not support only one function, so its connectivity shouldn’t be over interpreted). With the caveat that the connectivity differences observed do not in any way point to a cause and effect relationship – that is we don’t know if these differences existed before the first heroin intake and caused it or they appeared after as a result of drug consumption, I think the paper is still is worth reading.
Reference: Zou F, Wu X, Zhai T, Lei Y, Shao Y, Jin X, Tan S, Wu B, Wang L, Yang Z (November 2015, Epub 17 Aug 2015). Abnormal resting-state functional connectivity of the nucleus accumbens in multi-year abstinent heroin addicts.Journal of Neuroscience Research, 93(11):1693-702. doi: 10.1002/jnr.23608.Article | FREE PDF
No idea why the prefrontal cortex neuron is Australian, but here you go. Cartoon made by me with free (to the best of my knowledge) clipart elements. Feel free to use to your heart’s content.
There have been literally thousands of pages published about the dopamine function(s). Dopamine, which made its stage debut as the “pleasure molecule”, is a chemical produced by some neurons in your brain that is vital to its functioning. It has been involved in virtually all types of behavior and most diseases, from pain to pleasure, from mating to addiction, from working-memory to decision-making, from autism to Parkinson’s, from depression to schizophrenia.
Here is another account about what dopamine really does in the brain. Schwartenbeck et al. (2015) trained 26 young adults to play a game in which they had to decide whether to accept an initial offer of small change or to wait for a more substantial offer. If they waited too long, they would lose everything. After that, the subjects played the game in the fMRI. The authors argue that their clever game allows segregation between previously known roles of dopamine, like salience or reward prediction.
As expected with most fMRI studies, a brain salad lit up (that is, your task activated many other structures in addition to your region of interest), which the authors address only very briefly. Instead, they focus on the timing of activation of their near and dear midbrain dopamine neurons, which they cannot detect directly in the scanner because their cluster is too small, so they infer their location by proxy. Anyway, after some glorious mental (and mathematical) gymnastics Schwartenbeck et al. (2015) conclude that
1) “humans perform hierarchical probabilistic Bayesian inference” (p. 3434) (i.e. “I don’t have a clue what’s going on here, so I’ll go with my gut instinct on this one”)and
2) dopamine discharges reflect the confidence in those inferences (i.e. “how sure am I that doing this is going to bring me goodies?”)
With the obvious caveat that the MRI doesn’t have the resolution to isolate the midbrain dopamine clusters and that these clusters refer to two very distinct population of dopamine neurons (ventral tegmental area and substantia nigra) with different physiological, topographical, and anatomical properties, and distinct connections, the study adds to the body of knowledge of “for the love of Berridge and Schultz, what the hell are you DOIN’, dopamine neuron?”.
Reference: Schwartenbeck, P., FitzGerald, T. H., Mathys, C., Dolan, R., & Friston K. (October 2015, Epub 23 July 2014). The Dopaminergic Midbrain Encodes the Expected Certainty about Desired Outcomes.Cerebral Cortex, 25:3434–3445, doi:10.1093/cercor/bhu159.Article + FREE PDF
Like many studies that fill in unknown gaps in the body of knowledge, the paper below may not attract attention, except from the people in their narrow field. So let’s give it a little attention.
Michels et al. (2015) sought to map out the brain network underlying the control of urination using an fMRI scanner. They got 22 healthy adult males and they gave them furosemide, which is a diuretic, and then asked them to drink as much water as they want until they need to urinate. During this, “a condom catheter was attached to the penis of each subject” (p. 3370), to monitor the urine flow while in scanner. Then the testing would not start, oh no. The subjects were then submitted to an ultrasound to make sure the bladder was full. Then they were asked again how much they really needed to go pee. Then they go in the scanner in a supine position, where they are told to wait, then to imagine the starting of urination (but don’t pee just yet!), and finally, finally allowed to urinate. But then, cruelly, told to stop only 3 second into the act. And then the scanner cycle would repeat. Their champion peers (I cannot avoid the pun, I’m sorry) managed to pee 15 times in the scanner. I wonder how many subjects peed sans cue… (authors don’t mention that).
Fig. 1 from Michels et al. (2015) depicting the fMRI scan paradigm, which consisted of 2 randomly alternating blocks. SDV = strong desire to void.
Amazingly, under these conditions, there were seven men who could not urinate in the scanner. Authors call these non-voiders, or, as we commonly know them, the shy bladders or the bashful kidneys. Not surprisingly, non-voiders had lower activity in the pontine micturition center (PMC), a brain area, which, as its name implies, is responsible for urination. Also not surprisingly – for me at least, the authors find this interesting -, the non-voiders showed increased activity in the anterior midcingulate cortex (aMCC), which is an area involved in control. I guess you need some steely control to not pee after all that. The aMCC inhibits the urination-facilitation brain regions, such as the PMC.
Anyway, the main finding of the study is a detailed map of micturition supraspinal mechanisms, which consists of a slew of structures, each with its own function. I had never known how complicated peeing and not-peeing are until I read this paper. Jokes and cringes aside, this study is a welcome addition to understanding how we control our bodily functions and where to start looking when this control fails, shedding new light on the interplay between reflex and control.
Reference: Michels, L., Blok, B. F., Gregorini, F., Kurz, M., Schurch, B., Kessler, T. M., Kollias, S., & Mehnert, U. (October 2015, Epub Jun 26 2014). Supraspinal Control of Urine Storage and Micturition in Men-An fMRI Study. Cerebral Cortex, 25(10): 3369-80. doi: 10.1093/cercor/bhu140. Article |FREE FULLTEXT PDF
Drosophila melanogaster image illustrating sexual dimorphism and mating behavior. Credit: TheAlphaWolf (Wikimedia Commons)
Did you know that flies sing? True to the dictum that I just made up – ‘where is song, there is lust’ – it turns out not only that flies can sing, but they even have courtship songs! Granted, since they don’t have a larynx, the male flies sing by vibrating their wings in a certain way, which is unique to each fly species, and females listen with the feather-looking bit on top of their antennae, called arista. The behavior has generated enough research that a fairly hefty review about it has been published two years ago in Nature Reviews Neuroscience, pointing to a gene central to the male courtship circuitry and expressed only in the fly’s neurons, the fru gene (I bet it was called that way because when you make mutants you get fru–/fru– …).
Zhou et al. (2015) used a series of complicated experiments to successively activate or inhibit the neurons which express the fru gene, in order to identify the neural circuitry underlying hearing and processing the courtship songs. This circuitry is different in males and females, which makes sense since the serenading male expects different behaviors from his audience, depending on their sex; the listening males hurry to compete for the intended female and the females slow down and… listen carefully. Mind wondering: if I was the one serenading, wouldn’t I want to drive away the competitors, instead of drawing them in towards the object of my desire? Perhaps I want the competitors to also engage in courtship behavior so I can show off my wing vibrating prowess… Anyway, digression aside, in addition to figuring out which neuron does what, the authors managed to elicit courtship behavior in the listening males by optogenetically stimulating the 3rd and 4th order neurons in the newly identified circuit.
Besides being strangely interesting in itself, the research fills a gap in the understanding how courtship behavior is recognized, at least in fruit flies, which may be very useful information for other species as well, humans included.
Reference: Zhou, C., Franconville, R., Vaughan, A. G., Robinett, C. C., Jayaraman, V., & Baker, B. S. (21 September 2015). Central neural circuitry mediating courtship song perception in male Drosophila. Elife, 4:1-15. doi: 10.7554/eLife.08477. Article +FREE PDF
For the interested specialist, the MATLAB source code for analyzing calcium-imaging data can be found here.










 Most of the time the brain receives information from different senses about the same object or event. For example, to localize an object that makes noise we use both visual and auditory information – if these are available to us -, process called multisensory information integration (MSII).
Most of the time the brain receives information from different senses about the same object or event. For example, to localize an object that makes noise we use both visual and auditory information – if these are available to us -, process called multisensory information integration (MSII).

 When you’re sick you also feel awful: no appetite, weak, sleepy, feverish, achy, and so on. This is called, appropriately so, the sickness syndrome.
When you’re sick you also feel awful: no appetite, weak, sleepy, feverish, achy, and so on. This is called, appropriately so, the sickness syndrome.